Митохондрии, ЦТК, дыхательная цепь
Каждый норовит вдохнуть кислород, а выдохнуть - углекислый газ!
(народная мудрость)
И снова о митохондриях и о цикле Кребса
Это сложная тема, поэтому я возвращаюсь к ней снова и снова. Митохондрии в клетке выполняют много разных функций, и одной из важнейших будет аэробное окисление субстратов с помощью молекулярного кислорода, при этом создается электрический трансмембранный потенциал и запасается энергия (в виде АТФ). Важный нюанс заключается в том, что различно не только количество митохондрий в клетках разных типов, но и специализироваться они могут на разных функциях: в жировой ткани - на теплорегуляции, в нейронах - на генерации электрического потенциала, в мышцах - на синтезе АТФ, в печени - на метаболизме карбоновых кислот, и т.д. Это не значит, что остальные функции в митохондриях этих клеток отсутствуют. Это значит, что какая-то из них - доминирует [2].
Принято считать, что митохондрии имеют продолговатую форму. Однако Н.Л. Векшин в своих работах и в недавно вышедшей монографии [2], обобщающей все полученные им результаты, пишет, что не только. Шаровидная форма тоже распространена, и встречается чаще эллипсоидной. Митохондрии имеют свою ДНК, она имеет вес примерно 16 кДа и кольцевую форму. Поскольку митохондрии не умеют исправлять накапливающиеся ошибки в своей ДНК, то "плохие" ДНК утилизируются, а для кодирования нужных белков используется "нормальная" копия ДНК (всего в митохондрии имеется до 10 копий).
Интересно, что не менее 75% белков митохондрий (включая субъединицы ферментных комплексов) кодируются в ядре клетки и синтезируются в цитоплазме, а затем перетаскиваются в митохондрии белками-шаперонами. Это означает, что несмотря на то, что митохондриальная ДНК всегда передается по женской линии, тем не менее митохондриальная белковая "начинка" как минимум на 75% наследуется от обоих родителей.
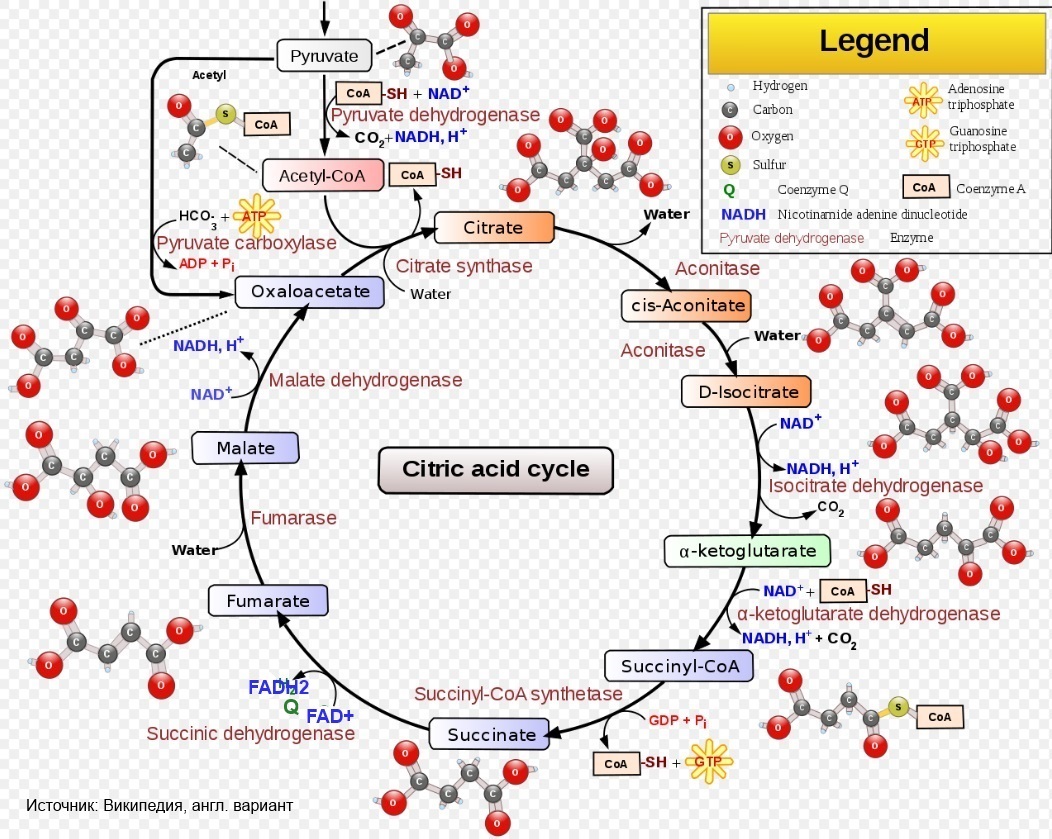
Д.Нельсон в замечательной работе (которую рекомендую каждому, кто хочет систематизировать свои знания по биохимии) "Основы биохимии Ленинджера" [1] пишет о том, что есть такое явление, как тунеллирование субстратов. Это означает примерно следующее: если пируват был перенесен в митохондрию и попал в цепкие лапы пируват-дегидрогеназного комплекса, а получившийся ацетил-КоА сконденсировался с оксалоацетатом, то дальше все пойдет по известной всем заинтересованным людям схеме (см. рис., взятый из Википедии).
Начинаем мы подробно разглядывать эту схему и видим, что в цикле Кребса происходит следующее:
- 2 раза - декарбоксилирование (уходит углерод в виде углекислого газа):
- с изоцитрата;
- с альфа-кетоглутарата;
- 4 раза - дегидрирование (уходит водород 3 раза в виде протона и восстановленного NAD, 1 раз - в виде восстановленного FAD* :
- с изоцитрата;
- с альфа-кетоглутарата;
- с сукцината;
- с малата.
* так нарисовано на схеме, но это вовсе не означает, что именно так оно происходит на самом деле.
Ну и CO2 выводится, восстановленные NAD и FAD окисляются обратно (кстати, где именно?), в дыхательную цепь идут электроны (кстати, откуда они вдруг берутся?), а субстраты переходят друг в друга и гоняются себе по кругу. Такое, по крайней мере, создается впечатление, когда читаешь книжку и разглядываешь схему. И как-то забываешь, что где-то там еще болтались протоны (кстати, где они?).
А потом начинаешь изучать поподробнее устройство митохондрий, и выплывают следующие подтвержденные исследованиями факты [2]:
- Внешняя мембрана митохондрий проницаема для мелких молекул (до 5 кДа);
- Внутренняя мембрана плохо проницаема для протонов и NADH, однако проницаема для сукцината (и малата) и прекрасно проницаема для кислорода;
- В матриксе митохондрий почти нет протонов;
- Дыхательная цепь - это не какая-то одна специальная структура, а несколько параллельно работающих трансмембранных ферментных комплексов;
- FAD не гуляет по матриксу, а встроен в дыхательную цепь (а именно - в специализированные ферментные комплексы. Если NAD и NADH свободно болтаются повсюду, то флавины нигде не болтаются, и более того: выход флавинов из соответствующих комплексов ассоциируется с повреждением дыхательной цепи и избыточным синтезом супероксида [2].);
- Сукцинат дегидрируется на кристах, а не в матриксе (то есть именно в дыхательной цепи специализированным ферментным комплексом, отсюда передача электронов на FAD, а не NAD);
- Убихинон (он же - коэнзим Q) "сидит" во внутренней мембране, цитохром C "плавает" между внутренней и внешней мембраной, а дегидрогеназные комплексы кроме всего прочего работают как помпы, выкачивая из матрикса протоны в межмембранное пространство, где они будут соединяться с кислородом на цитохромоксидазе и образовывать воду. При этом убихинона гораздо больше, чем дегидрогеназ и цитохромов - просто потому, что иначе дыхательная цепь не сможет быстро работать;
- "Круга" не существует, это всего лишь модель, иллюстрирующая последовательность превращений, а фактически субстраты находятся кто где и диффундируют туда-сюда, насколько позволяют условия.
Дыхательная цепь
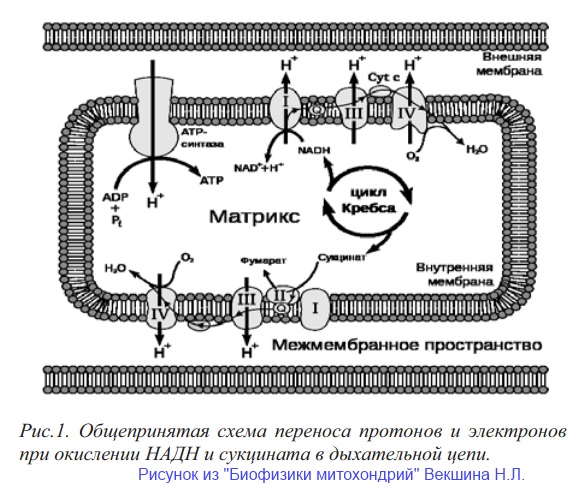
То, что мы называем для простоты дыхательной цепью, на самом деле представляет собой систему встроенных во внутреннюю мембрану ферментных комплексов. При этом внутренняя мембрана сама по себе состоит из белков и липидов в соотношении 3:1, а вовсе не является обычным липидным бислоем, как это принято считать (и если учесть относительный размер белков и фосфолипидов - это скорее белковый слой с небольшой липидной составляющей) [2]. Ферментные комплексы дыхательной цепи включают в себя:
- NADH-дегидрогеназный комплекс;
- b-c-цитохромный комплекс;
- цитохромоксидазный комплекс;
- сукцинатдегидрогеназный комплекс.
На первом шаге работает NADH-дегидрогеназный комплекс. Этот комплекс частично встроен в мембрану, а частично торчит из нее. Он содержит железо-серные кластеры, а также какой-то из флавинов (считается, что это ФМН, но это не точно - возможно, что это ФАД). Комплекс окисляет NADH из матрикса, при этом окисление может происходить двумя путями:
- два электрона отдаются на кластеры, и кластеры уже восстанавливают убихинон, отдавая ему 2 электрона;
- один гидрид-ион отдается на флавин или убихинон (он же - коэнзим Q). Если гидрид-ион передается на флавин, то флавин уже восстанавливает убихинон.
В любом случае получается восстановленный убихинон (то есть убихиноЛ), но его формы чуть различаются в зависимости от путей окисления.
Сукцинатдегидрогеназный комплекс тоже содержит флавопротеин и железосерный белок. Этот комплекс окисляет сукцинат до фумарата, и при этом конечным акцептором электронов является все тот же убихинон (но не всегда). А ФАД, нарисованный на картинке, изображающей ЦТК, - это промежуточное звено комплекса, то есть часть фермента. Он не является самостоятельной единицей и не таскает электроны туда-сюда, перемещаясь сам, а перебрасывает их, как теннисист отбивает мяч ракеткой. Ну и интересной особенностью сукцинатдегидрогеназы является то, что она кодируется ядерной, а не митохондриальной ДНК.
Это надо отметить отдельно: сукцинатдегидрогеназа, во-первых, есть только в митохондриях, а во-вторых, она находится именно в дыхательной цепи. И кодируется при этом ядерной ДНК! [2].
Цитохромоксидаза также является металлоферментом и содержит в своих активных центрах как железо, так и медь. Считается, что этот ферментный комплекс последовательно окисляет 4 молекулы цитохрома С, при этом принимает 4 электрона и восстанавливает молекулярный кислород O2 до H2O, образуя, соответственно, две молекулы воды на каждую молекулу кислорода. Что касается протонов, то не факт, что их в реакции участвует только 4 - может быть, и больше, так как, возможно, образуется протонный градиент, а для этого нужно, чтобы перекачивалось больше протонов в межмембранное пространство (но это не точно).
В популярной литературе, когда говорят об антиоксидантах, упоминают витамин С (аскорбиновую кислоту) и коэнзим Q10 (убихинон). Антиоксиданты - это вещества, способные служить прямыми донорами электронов (то есть окисляться) для других веществ (находящихся в окисленной форме) и тем самым восстанавливать эти окисленные вещества (то есть возвращать им недостающие электроны). Это справедливо и для работы цитохромоксидазы (в лице находящихся в ней ионов железа и меди), правда, с маленьким нюансом: если эти доноры окажутся в нужное время в нужном месте и в нужной форме.
А теперь посмотрим на цикл Кребса следующим образом. Оксалоацетат и малат - это, можно сказать, близнецы-братья, отличающиеся только по степени окисленности-восстановленности (редокс-статусу). Малат и фумарат тоже переходят друг в друга, была бы вода. А вот фумарат из сукцината образуется в дыхательной цепи, и это существенная трансформация. Так же, как и образование сукцината из альфа-кетоглутарата (через сукцинил-КоА). А альфа-кетоглутарат при этом может получаться из глутамата посредством транс- или дезаминирования.
Малат, как мы знаем, может свободно покидать митохондрии для, например, глюконеогенеза (кстати, при помощи какого транспортера?). Альфа-кетоглутарат может попадать в митохондрии (или образовываться там из глутамина-глутамата? Этот момент надо уточнить). Таким образом вполне может существовать не полный цикл Кребса, а лишь его часть: от альфа-кетоглутарата до малата. Можно даже посчитать, сколько именно АТФ дает такой "урезанный" цикл. Не совсем, правда, понятно, как тогда быть с туннелированием субстратов, про которое писал Нельсон [1] (думаю, что исследователи метаболизма раковых клеток этот вопрос изучали, так что нужно только найти литературу).
Но самое интересное здесь то, что, возможно, именно так функционируют митохондрии в клетках волосяного фолликула. А если они вдруг по какой-то причине начнут функционировать по-другому, то фолликул не будет получать того, что ему нужно, а станет получать то, что ему не нужно, со всеми вытекающими из этого последствиями в виде ухода в телоген, миниатюризации, а то и драматичнее**.
** например, при химиотерапии онкологических заболеваний. В метаболизме раковых клеток и в метаболизме волосяного фолликула есть много общего, а значит те средства, которые воздействуют на метаболизм раковых клеток, будут так же влиять и на метаболизм клеток фолликула - и вызывать потерю волос. Это логично, и остается только рассчитывать на то, что ученые что-нибудь придумают по этому поводу (пока что кое-где используют метод охлаждения скальпа, и он более-менее спасает фолликулы. Но доступен, видимо, не везде).
Продолжение следует.
Литература:
-
Д. Нельсон
Основы биохимии Ленинджера, том 2 "Биоэнергетика и метаболизм"
Издательство "Бином. Лаборатория знаний", 2014 -
Н.Л. Векшин
Биофизика митохондрий
ООО "Фотон век", 2019