О метаболизме волосяного фолликула: глутаминолиз и аэробный гликолиз
В последнее время складывается впечатление, что основное внимание ученых привлечено к сигнальным путям регуляции процессов жизнедеятельности волосяного фолликула, в то время как фундаментальный вопрос о том, как и чем обеспечивается энергетический баланс в этом удивительном органе, остается вне поля внимания. Может быть так получается потому, что все давным давно в этом вопросе разобрались. Но может оказаться, что просто забыли. Если первое, то повторить еще раз известные всем факты все же будет полезно: повторенье - мать ученья. Ну а если второе, то тем более будет полезно. Ибо работы в этом направлении велись ни много, ни мало, а в прошлом веке, на волне подъема биохимии: не зря исследователи ссылаются на работы товарища Кребса, а не просто что-то говорят о цикле трикарбоновых кислот и его интермедиатах. Одна из первых (относительно доступных) статей [3] на эту тему датируется 1968 годом, и до сих пор опубликовавший ее журнал хочет за нее свои 25 долларов!
В любом случае обобщим то, что мы знаем про метаболизм волосяного фолликула и затем посмотрим на эти данные с учетом того, что накопали ученые исследователи про другие ткани и органы, отличающиеся высокой скоростью деления клеток.
Вкратце суть метаболизма волосяного фолликула сводится к следующим метаболическим путям:
- Гликолиз
- Образование лактата из пирувата
- Пентозофосфатный путь
- Цикл трикарбоновых кислот (он же - цикл Кребса)
- Образование альфа-кетоглутарата из глутамина
- Возможно - синтез гликогена
- Возможно - глюконеогенез из лактата, то есть свой собственный локальный цикл Кори
Понятно, что измерить все биохимические параметры in vivo довольно сложно, поэтому исследования проводятся in vitro, при этом придумываются разные хитрые способы, чтобы максимально приблизить условия "в пробирке" к условиям "вживую". Если волос просто выдрать, то метаболизм полученного фолликула может быть нарушен стрессом от выдирания. Однако исследователи Уильямс, Филпотт и Кили [1] придумали способ, как изолировать волосяные фолликулы, не нарушая их жизнедеятельности. Такие фолликулы способны нормально продуцировать волос до 10 дней, поэтому модель оказалась вполне репрезентативной. Именно на такой модели было выяснено, что конкретно является топливом для фолликула, а что - не является.
Предсказуемо оказалось, что основным поставщиком энергии для волосяных фолликулов является глюкоза. Именно распад глюкозы дает основное количество АТФ для жизнедеятельности фолликула. Количественно энергетическая емкость тканей характеризуется соотношением Аткинсона: АТФ/АМФ; для волосяных фолликулов в фазе анагена это соотношение составляло в среднем 7.0, что дает энергетическую емкость примерно 0.81+-0.08. Глюкоза поступает в клетки фолликулов посредством транпортеров GLUT1 [5] и дальше утилизируется (а то, что не утилизируется, складывается в виде гликогена? Похоже, что так [4], [5].)
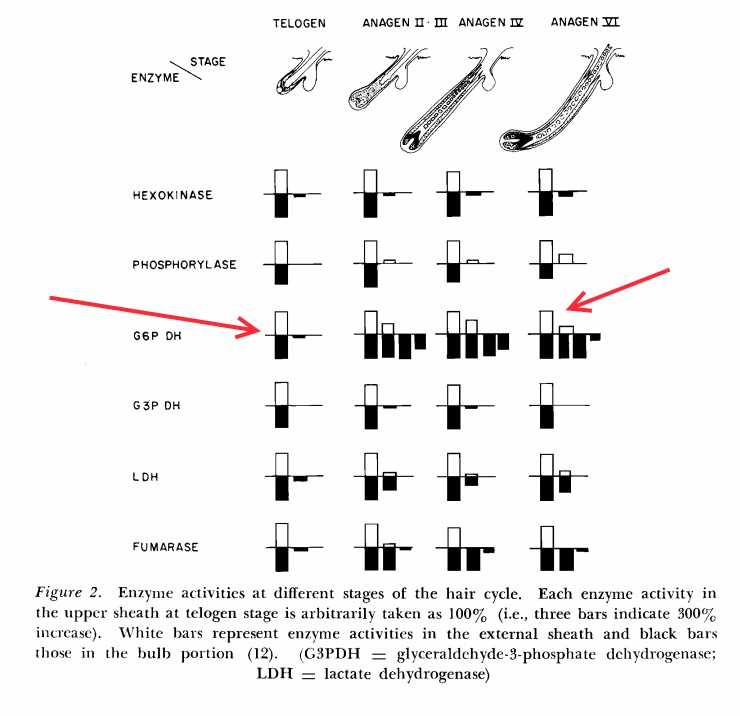
Глюкоза, поступившая в клетки, запирается в них через фосфорилирование, образуя глюкозо-6-фосфат (при этом активность гексокиназы в анагене увеличивается лишь незначительно в клетках матрикса и остается постоянной в клетках внешней корневой оболочки [2], см. рисунок. И это, очевидно, означает, что фолликул потребляет почти одно и то же количесво глюкозы вне зависимости от того, растет он или отдыхает). Дальше бОльшая часть глюкозы распадается до пирувата, бОльшая часть которого в свою очередь восстанавливается до лактата. А часть пирувата все-таки попадает в цикл Кребса, но это не слишком большая часть утилизованной глюкозы. Таким образом волосяной фолликул основную часть АТФ получает посредством аэробного гликолиза, причем это справедливо как для анагеновых, так и для телогеновых фолликулов [2]. А этот факт означает, что фолликулу нужно не так много кислорода, поскольку лишь небольшая часть электронов попадает на дыхательную цепь и нуждается в связывании с кислородом воздуха (отсюда вопрос о роли кровоснабжения: практика показывает, что оно необходимо, однако экспериментально полученные данные говорят скорее о том, что его основная роль - снабжение фолликулов питательными веществами и уже в меньшей степени - кислородом).
Чем же тогда отличается метаболизм анагеновых фолликулов от метаболизма фолликулов в телогене? Тем, что в анагеновых фолликулах часть глюкозо-6-фосфата отправляется на образование пентоз, а в телогеновых фолликулах пентозофосфатный путь неактивен [2]. В анагене резко увеличивается активность ключевого фермента пентозофосфатного пути - глюкозо-6-фосфат-дегидрогеназы (см.рис.), причем происходит это именно в клетках нижней части фолликула, в то время как в клетках внешней корневой оболочки активность этого фермента тоже увеличивается, но далеко не столь драматично. Это тоже вполне предсказуемо и объяснимо с точки зрения здравого смысла: активно делящимся клеткам матрикса очень нужны пентозы и/или NADPH для синтеза нуклеиновых кислот.
Почему же тогда в анагене продолжается преимущественное преобразование пирувата в лактат, причем происходит это несмотря на наличие нормальных митохондрий? Возможно, дело в соотношении кофакторов NAD+ и NADH, а также NADP+ и NADPH. Но только ли в этом?
Вспомним: для гликолиза нужен NAD+ (при окислении глицеральдегид-3-фосфата). Для того, чтоб пируват вошел в цикл Кребса, нужен NAD+. А при активном гликолизе накапливается NADH и становится все меньше NAD+. Восстановление пирувата до лактата - это самый простой способ получить из NADH такой необходимый NAD+. А лактат затем отправляется окисляться обратно до пирувата, обычно это делается в печени, и называется этот процесс циклом Кори [8]. Однако вопрос: только ли этим ограничивается функция лактата? Не слишком ли дорого - отправлять весь этот лактат в печень? Может, у фолликула есть свой собственныймаленький свечной заводикспособ конвертировать лактат обратно в пируват? А если есть, то опять же, зачем? Мы ведь помним, что гликолиз и глюконеогенез взаимно исключают друг друга? Вопросов так много, а ответ может прийти, откуда не ждали.
Глутамин - допинг для фолликула
Глюкоза не является единственным топливом, необходимым для оптимальной жизнедеятельности волосяного фолликула. Оказалось, что наилучшие результаты, выражающиеся в максимальном росте волоса, были получены, когда топливом являлась смесь глюкозы и глутамина в пропорции 2мМ глутамина на 11.1 мМ глюкозы (1.64+-0.10 мм за 6 дней). В том случае, когда топливом была одна глюкоза, рост волоса составил всего лишь 0.38+-0.13 мм за 6 дней. Аналогичный результат получен и для одного глутамина (без глюкозы).
Интересно, что если в растворе с глутамином заменить глюкозу на бета-гидроксибутират или пальмитат, то результат сразу резко снизится: в первом случае до 0.52+-0.11 мм, а во втором 0.34+-0.08 мм за 6 дней. Если заменять не глюкозу, а глутамин, то получим соответственно 1.07+-0.08 мм и 0.68+-0.03 мм. То есть жирные кислоты не способны адекватно выполнять те функции, которые выполняют глюкоза и глутамин.
Что такого делает глутамин, почему он в сочетании с глюкозой обеспечивает максимальное деление клеток? Использование глутамина снижает необходимость оксидативного использования глюкозы (и в волосяных фолликулах это тоже так: использование глутамина снижает потребление глюкозы в принципе [1]). Снижение активности цикла Кребса понижает вероятность утечки электронов и свободных радикалов, значит, снижается вероятность ошибок при делении клеток. В целом получается очень интересная картина: координированное использование глюкозы и глутамина плюс производство большого количества лактата напоминает паттерны, характерные для злокачественных опухолей [6] (эти паттерны широко изучаются в последнее время). И дело не только в эффекте Варбурга и не только в глутаминолизе (помним, что альфа-кетоглутарат - это важный кофактор реакций метилирования гистонов), но и в самой продукции лактата: может быть, именно повышение активности ЛДГ и рост количества лактата есть ключ к иммунной привилегии волосяного фолликула [7]?
Это непаханное поле для дальнейших исследований. Возможно, кто-то этим уже занимается. Что ж, проследим.
Продолжение следует.
Литература:
-
Rebecca Williams, Michael P. Philpott, Terence Kealey
Metabolism of freshly isolated human hair follicles capable of hair elongation: a glutaminolytic, aerobic glycolytic tissue
1993 -
Kenji Adachi, Susumu Takayasu, Iwao Takashima, Motonari Kano, Shigeo Kondo
Human hair follicles: metabolism and control mechanisms
1970 -
Kenji Adachi, Hideo Uno
Glucose metabolism of growing and resting human hair follicles
1968
Доступен только абстракт, а вообще за статью хотят денег; но все неплохо изложено в [2] -
Kuniaki Adachi, Yasusi Watanabe, Kuniyo Inouye
Activity of Glucose-6-phosphate 1-Dehydrogenase in hair follicles with male-pattern alopecia
1999 -
Katarzyna Figlak, Ralf Paus, Mike Philpott
Glycogen metabolism and Cori cycle in human hair follicles
2017 -
Chiara Damiani, Riccardo Colombo, Daniela Gaglio, Fabrizia Mastroianni, Dario Pescini, Hans Victor Westerhoff, Giancarlo Mauri, Marco Vanoni, Lilia Alberghina
A metabolic core model elucidates how enhanced utilization of glucose and glutamine, with enhanced glutamine-dependent lactate production, promotes cancer cell growth: The WarburQ effect
2017 -
Almut Brand, Katrin Singer, Gudrun E. Koehl, Wolfgang Mueller-Klieser, Kathrin Renner, Marina Kreutz
LDHA-Associated Lactic Acid Production Blunts Tumor Immunosurveillance by T and NK Cells
2016 -
Д. Нельсон
Основы биохимии Ленинджера, том 2 "Биоэнергетика и метаболизм"
Издательство "Бином. Лаборатория знаний", 2014